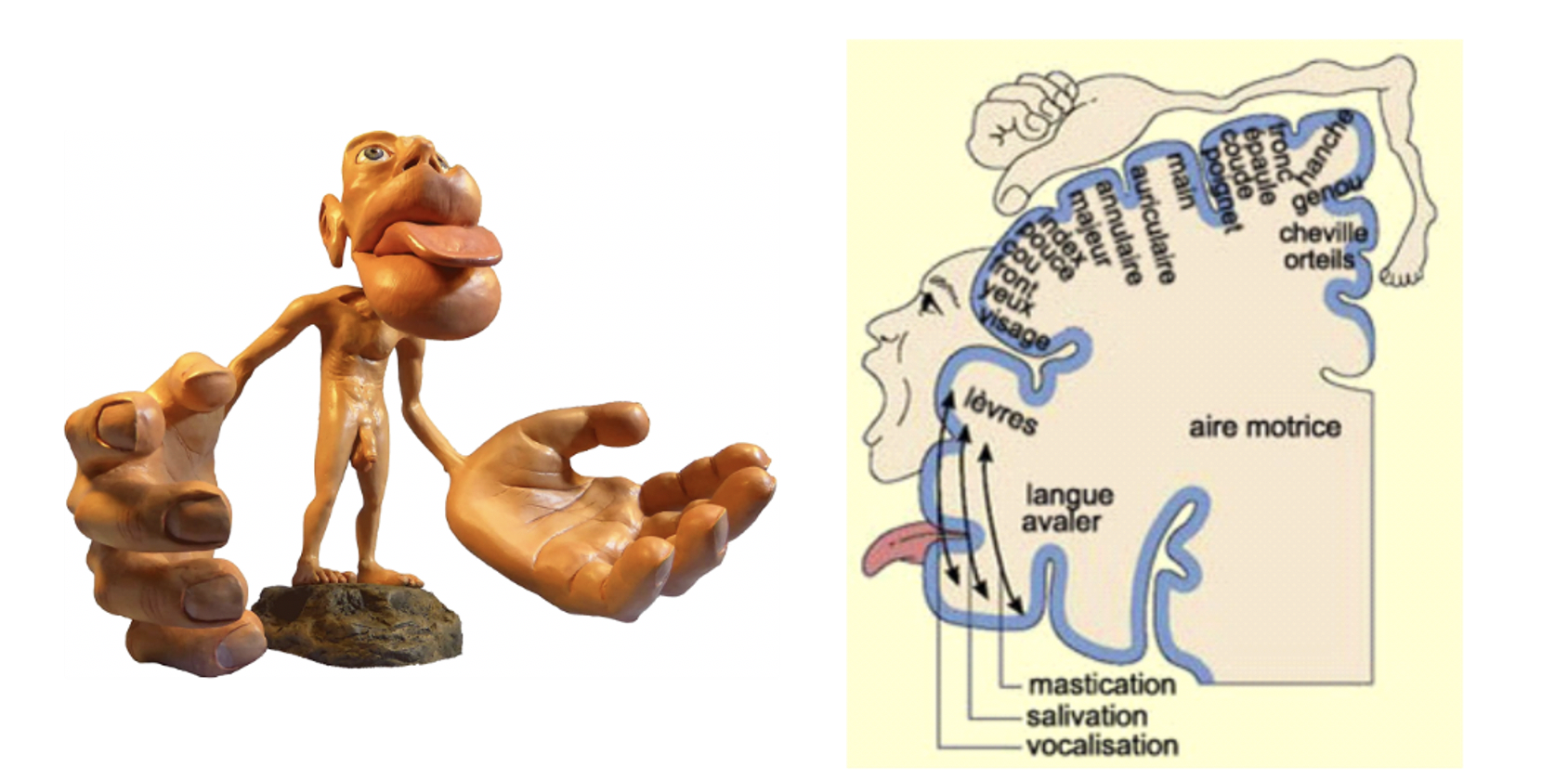
Figure 1 – L’homoncule de Penfield
La principale interprétation dérivant de l’homoncule fut de
supposer un lien entre l'étendue corticale et fonction. Autrement dit, si le cerveau alloue beaucoup d’espace à une fonction, c’est que cette fonction doit être particulièrement importante et développée. Suivant cette logique, les cerveaux d’individus aux fonctions sensorimotrices exceptionnelles devraient en porter les stigmates. Dans cette optique il apparait pertinent d’
étudier les cerveaux d’athlètes.
En 2014, deux chercheurs japonais ont mesuré l’activité cérébrale du cerveau du joueur de football Neymar Jr. lorsqu’il effectuait des mouvements de pieds (Naito and Hirose, 2014). De manière surprenante, la comparaison de ses activités à celles d’individus contrôles (des athlètes pratiquant un sport dont les mouvements des pieds sont moins importants que dans le football ou des footballeurs amateurs) mit en évidence que l’étendue spatiale des activités du cerveau de Neymar était bien plus restreinte (Figure 2). Ce résultat semble paradoxal : comment Neymar peut-il être si performant avec ses pieds si ce rôle est porté par une toute petite partie de son cerveau ? Et si l’important n’était pas l’étendue ni la localisation mais l’efficacité des traitements neuronaux ?
“Comment Neymar peut-il être si performant avec ses pieds si ce rôle est porté par une toute petite partie de son cerveau ? Et si l’important n’était pas l’étendue ni la localisation mais l’efficacité des traitements neuronaux ? ”
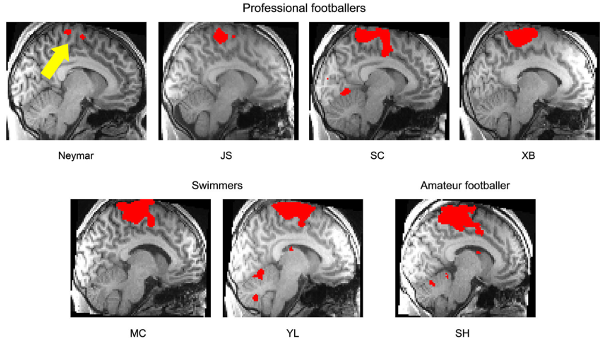
Figure F2 – Activités des aires motrices lors de mouvements du pied
Activités mesurées en imagerie fonctionnelle chez plusieurs footballeurs professionnels dont Neymar Jr., des nageurs et un footballeur amateur dans une tâche leur demandant d’effectuer des mouvements isolés du pied droit à faible amplitude à une fréquence de 1Hz (1 mouvement par seconde). L’étendue des activités (rouge) mesurées au niveau des aires motrices primaires est inversement corrélée au degré d’expertise des mouvements de pieds chez ces sujets (Naito and Hirose, 2014).
Le développement récent de l’application de cette hypothèse (en anglais, Neural Efficiency Hypothesis, NEH) aux athlètes apporte des éléments théoriques, conceptuels et pratiques. En particulier l’entrainement sportif semble engendrer des phénomènes d’adaptation du fonctionnement cérébral (neuroplasticité fonctionnelle) dont la conséquence est l’optimisation de l’utilisation des ressources cérébrales allouées à la réalisation des tâches qui sont répétées. La compréhension des mécanismes sous-jacents et du décours temporel de cette neuroplasticité constituerait une avancée considérable des perspectives d’amélioration des performances des athlètes.
Une revue systématique récente (Li and Smith, 2021) a dénombré 28 études soutenant la NEH chez les athlètes. Ces études convergent sur l’observation que les athlètes de haut niveau réalisent les tâches liées à leur sport de manière plus performante (plus rapidement, plus précisément, en commettant moins d’erreurs) tout en leur allouant moins de ressources cérébrales. La généralisation de cette observation à travers différentes disciplines sportives et des contextes dans lesquelles les actions sont soit guidées par des stimuli externes soit par des traitements internes témoigne du rôle fondamental de l’entrainement de haut niveau dans la mise en place de l’efficacité neuronale (Table 1).
“Les athlètes de haut niveau réalisent les tâches liées à leur sport de manière plus performante tout en leur allouant moins de ressources cérébrales.”

Table 1 – Dénombrement du nombre d’études dont les résultats obtenus soutiennent l’hypothèse de l’efficacité neuronale chez l’athlète dans diverses disciplines sportives (Li and Smith, 2021)
Plus précisément, les potentiels évoqués moteurs au niveau de l’aire motrice primaire des athlètes sont évoqués plus rapidement et sont de plus faibles amplitudes. Ces potentiels constituent le signal de sortie de la commande motrice au niveau du cortex avant d’être relayé par les motoneurones de la moelle épinière pour parvenir jusqu’à la jonction neuromusculaire et provoquer le recrutement des groupes musculaires cibles. Ainsi les signaux moteurs générés par les cerveaux des athlètes, bien que quantitativement moins importants que ceux des non-athlètes parviennent à soutenir des réponses motrices plus performantes. Ces résultats électrophysiologiques sont appuyés par l’observation en imagerie fonctionnelle de la diminution des activités neuronales au sein des principales régions du système moteur. De manière intéressante, ces observations ne sont pas confinées au système moteur mais s’étendent aux régions impliquées dans les traitements visuo-spatiaux, dont dépendent largement les performances des actions sportives étudiées. Ceci est cohérent avec la NEH qui ne s’applique pas seulement aux traitements moteurs mais plus généralement à tout traitement qui peut être répété, appris, automatisé. Ainsi au cours de l’apprentissage sportif, des changements structurels et fonctionnels permettent d’optimiser les traitements et de réduire la consommation en ressources.
Ce versant ne constitue cependant qu’une partie du prisme. En effet ces observations furent réalisées dans des contextes sportifs relativement peu complexes demandant de traiter peu d’informations et dont le degré d’abstraction des décisions est faible. En dehors du sport, la NEH est à la fois soutenue et contestée par des résultats apparemment contradictoires obtenus chez des individus ayant acquis une expertise cognitivo-motrice. Ces résultats soutiennent en réalité une dualité de processus mis en jeu. Dans des situations peu complexes, les experts sont plus performants tout en déployant moins de ressources attentionnelles et cérébrales. En revanche, lorsque les situations deviennent complexes et/ou inattendues, les experts sont capables d’augmenter leurs capacités attentionnelles, de traitements et de contrôle cognitif, supportées par l’activation cohérente d’ensembles d’aires corticales, principalement fronto-pariétales. Ainsi le régime de fonctionnement de leurs activités cérébrales s’adapte flexiblement au contexte décisionnel. Par rapport aux non-experts, cette flexibilité est plus rapide à transiter entre les deux modes, plus marquée en termes de différences d’activités et davantage adaptée aux besoins contextuels. De manière analogique, on peut considérer que les activités neuronales des athlètes transitent entre optimisation automatique et optimisation contrôlée dépendamment de la demande pour maintenir un haut niveau de performance tout en optimisant l’utilisation de ressources cérébrales (Figure 3). L’entrainement ayant pour effet d’améliorer ces différentes optimisations y compris l’efficacité des transitions entre les différents modes de fonctionnements. Ces apprentissages sont supportés par des changements structurels et fonctionnels de l’organisation des réseaux corticaux mis en jeu lors de la réalisation des tâches apprises.
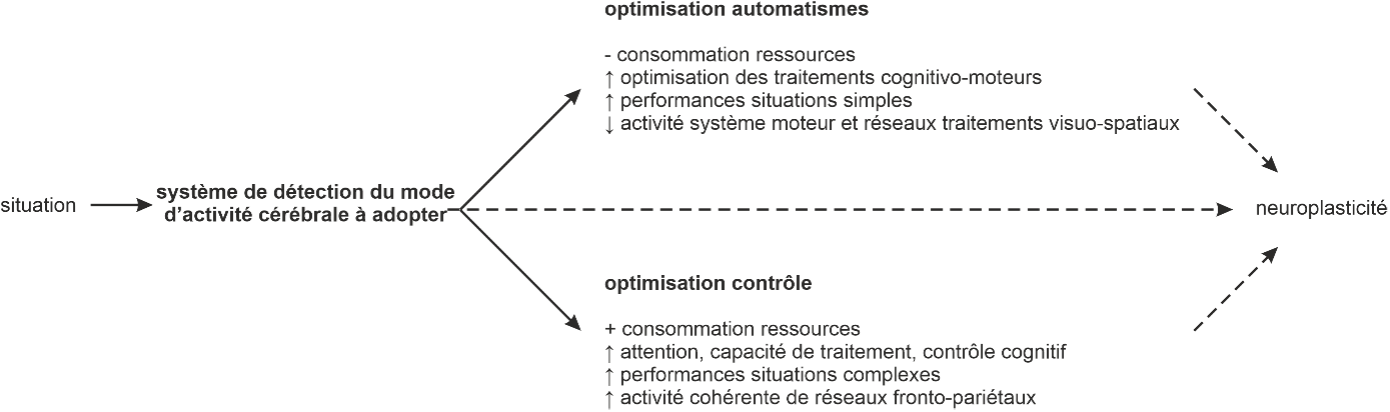
Figure 3 – Le cerveau d’athlète : une dualité d’optimisation des ressources cérébrales. Représentation schématique des hypothèses d’optimisation des ressources cérébrales chez l’athlète. Une situation rencontrée dans le domaine d’expertise de l’athlète va, dépendamment de sa complexité, soit déclencher la mise en route d’un mode de fonctionnement d’optimisation des automatismes, soit d’optimisation du contrôle. L’efficacité de ces modes de fonctionnement, tout comme celle du système de détection du mode à adopter sont améliorés lors de l’apprentissage grâce à une neuroplasticité structurelle et fonctionnelle des réseaux cérébraux impliqués.
Si le rôle d’un entrainement prolongé dans la mise en place des phénomènes d’optimisation des ressources cérébrales que nous venons de décrire est indiscutable, il réside de nombreuses inconnues quant à sa temporalité. Comment la neuroplasticité se met-elle en place dans le temps ? En d’autres termes combien de temps faut-il pour devenir un expert ? Y a-t-il des périodes de la vie plus prolifiques ?Il convient probablement mieux de raisonner en termes de nombre et de qualité de répétitions de situations similaires engendrant une restructuration des activités neuronales dans le temps. En effet dès quelques centaines de répétitions d’une tâche, par exemple manuelle, on peut observer l’augmentation des activités au niveau des représentations corticales motrices de la main chez l’adulte (Kami et al., 1995). En revanche les restructurations plus complexes observées chez les athlètes n’occurrent qu’après un entrainement prolongé dans le temps. Le cerveau a donc besoin à la fois de sollicitations répétées mais également de temps pour optimiser le traitement de ces demandes. En effet, le degré d’optimisation semble inversement corrélé à ces variables comme en témoigne l’étendue des régions motrices actives lors de mouvement du pied respectivement de plus en plus grandes chez Neymar, d’autres footballeurs professionnels, des sportifs de haut niveau non-footballeurs et des amateurs (Figure 2).
L’organisation du système nerveux central supporte des traitements modulaires. Cette structuration permet d’apprendre des associations, des concepts et de représenter des généralisations. Ainsi nous ne devons pas apprendre de nouveau chaque nouvelle tâche ou situation que nous rencontrons. Notre cerveau a la capacité de baser ces nouveaux apprentissages d’après nos expériences et connaissances acquises dans des contextes similaires. La question qui se pose c’est de savoir si on peut dégager une mesure de la similarité nécessaire entre plusieurs situations pour être capable de transférer les compétences acquises dans l’une vers une autre. De toute évidence des transferts de compétences ont lieu dans le domaine sportif. Par exemple les pratiquants de sports de raquettes sont capables d’apprendre plus rapidement un autre sport de raquettes par rapport à d’autres sportifs. Cela veut bien dire qu’il y a un transfert de compétences. Ces transferts sont cependant très limités aux exécutions motrices très similaires. La raison réside probablement dans le fait que l’apprentissage sensorimoteur lié à un sport est très précis et consiste en des séquences complexes d’actions. Ces séquences mettent en jeu des circuits neuronaux qui sont devenus spécialisés. De fait leur efficacité est difficilement transférable à d’autres situations ayant leur propre complexité sensorimotrice (Simonet et al., 2019).
Dans le cadre conceptuel de la NEH, on peut néanmoins se demander si certains éléments sont plus transférables que d’autres. L’automatisation dans les situations simples ? Le contrôle dans les situations complexes ? La capacité de détecter si une situation est simple ou complexe et d’allouer plus ou moins de ressources cognitives en conséquence ?
À l’heure actuelle, il n’y a pas de réponse claire a toutes ces questions. Néanmoins une hypothèse probable estime qu’il peut y avoir dans une certaine proportion transfert de ces différents traitements. En particulier les musiciens, danseurs et sportifs semblent apprendre plus rapidement à discriminer les situations complexes des simples et ce même en dehors de leur domaine d’expertise. Différents réseaux d’aires cérébrales supportent les phases successives de l’apprentissage. Un ensemble d’aires contribue particulièrement à la phase au début d’un apprentissage. Ces réseaux permettent de détecter les changements, les erreurs - et de les corriger -, mais aussi de déployer de l’attention et de la mémoire de travail. Les performances de ces réseaux semblent particulièrement efficaces chez les individus pratiquant une activité sensorimotrice complexe de manière répétée et continue comme le font les athlètes. En d’autres termes il semble possible d’entrainer les capacités de traitement de ces réseaux. Selon cette hypothèse, notre cerveau est donc capable d’apprendre à apprendre (Seidler, 2010).



